Research highlights|Meir, Yigal
Intracellular networks in bacteria
Bacteria are constantly sensing their environments and adjusting their behavior accordingly. Signaling occurs through networks of proteins and nucleic acids, culminating in changes of gene expression and so changes in the proteins of the cell. We are focused on the architecture of these intracellular networks. What is the relation between network architecture and function? For example, can we understand the selection of architectures in terms of general information-processing concepts such as signal to noise, memory, and adaptation? Even in a single bacterium such as E. coli, there are hundreds of coexisting networks. Our belief is that a deep study of a small number of "model" networks will yield general tools to analyze information processing by cell. It is important to choose these model networks carefully. The network components should be well characterized and the physiological function of the network should be known and subject to quantitative measurement. Probes of the internal dynamics of the network such as fluorescence resonance energy transfer (FRET) or direct imaging of dynamic spatial structure, will be critical in developing and testing quantitative models. It will also be important to choose networks which complement each other well, spanning a broad range of architectures and functions. A preliminary list includes (i) chemotaxis, which requires adaptation and rapid response to changing chemical concentrations, (ii) cell-division networks, where accuracy and checkpoints are essential, and (iii) metabolic networks which tie together diverse inputs to maintain homeostasis.
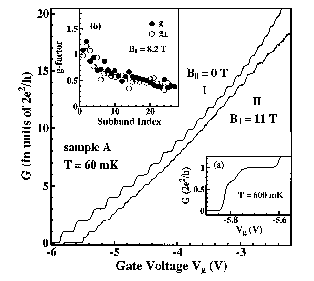
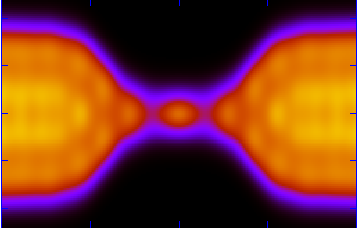
Solving the "0.7 Anomaly"
The simplest nanoelectronics device, and the basic building block for more complicated devices, is a "quantum point contact," a constriction connecting large electron reservoirs (see picture on the right). According to quantum mechanics the conductance through such a device should increase as the gap grew bigger by integer steps of universal value. Surprisingly, an additional first step approximately 0.7 times the expected universal value had also been observed, which became known as "the 0.7 anomaly" (see picture on the left). Previously we demonstrated that the phenomenology of this anomaly can be explained by the existence of a magnetic impurity, a localized electron in the quantum-point contact PRL 89, 196802 (2002). Using extensive density-functional calculations we demonstrated impurity.png the emergence of a magnetic impurity at the quantum point contact (see picture on the right) because a lower density of the electrons near the quantum point attracts the other electrons towards it. The wavy nature of such electrons then causes ripples, trapping an electron and causing the 0.7 anomaly.
 |
 |
 |
- Further reading:
- Magnetic impurity formation in quantum point contacts (Nature, 2006)
- Article is News and Views in Nature Physics
- Making the paper (Nature, 2006)